nukleobaser är byggstenarna från vilka de långa kedjorna av DNA- och RNA-molekyler är sammansatta i sin fosforylerade nukleotidform.
I DNA, som bildar dubbla strängar som liknar rep, bildar de fyra förekomna nukleiska baserna fasta parningar med respektive komplementär bas via vätebryggor. Nukleobaserna består av antingen ett bicykliskt purin eller ett monocykliskt pyrimidinskelett.
Vad är nukleobaser?
De fyra nukleobaserna adenin, guanin, cytosin och tymin, som byggstenar i de långa DNA-dubbla helixmolekylkedjorna, bildar de konstanta parningarna adenintymin (A-T) och guanin-cytosin (G-C).
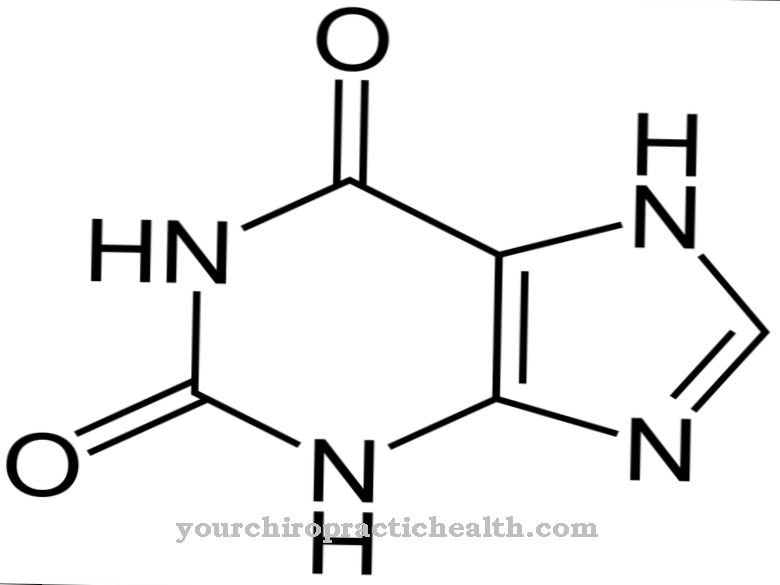
De två baserna adenin och guanin består vardera av en modifierad bicyklisk sex- och fem-ledarsring av den grundläggande purinstrukturen och benämnes därför också purinbaser. Grundstrukturen för de andra två nukleiska baserna, cytosin och tymin, består av en heterocyklisk aromatisk sexledad ring, vilket motsvarar ett modifierat pyrimidinskelett, varför de också kallas pyrimidinbaser. Eftersom RNA oftast finns som enstaka strängar finns det initialt inga baspar. Detta sker endast under replikering via mRNA (messenger RNA).
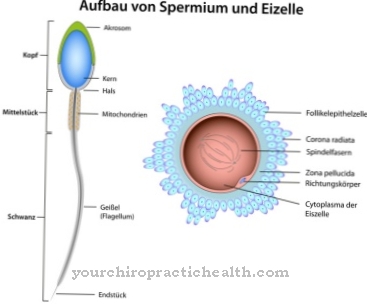
Kopian av RNA-strängen består av de komplementära nukleobaserna analoga med den andra DNA-strängen. Den enda skillnaden är att uracil ersätter tymin i RNA. DNA- och RNA-kedjemolekylerna bildas inte i sin rena form av nukleobaserna, utan snarare, i fallet med DNA, kombineras de med 5-socker deoxiribos för att bilda motsvarande nukleosid. För RNA består sockergruppen av ribos. Dessutom fosforyleras nukleosiderna med en fosfatrest för att bilda så kallade nukleotider.
Purinbaserna hypoxantin och xantin, som också förekommer i DNA och RNA, motsvarar modifierad tymin. Hypoxantin bildas från adenin genom att ersätta aminogruppen (-NH3) med en hydroxylgrupp (-OH), och xantin bildas från guanin. Båda nukleobaserna bidrar inte till överföring av genetisk information.
Funktion, effekt och uppgifter
En av de viktigaste funktionerna hos nukleobaserna, från vilka de dubbla DNA-strängarna är byggda, är att visa närvaro vid den avsedda positionen.
Sekvensen för nukleobaserna motsvarar den genetiska koden och definierar typen och sekvensen av aminosyror från vilka proteiner är sammansatta. Detta betyder att den viktigaste funktionen hos nukleobaserna som en del av DNA består av en passiv, statisk roll, dvs att de inte aktivt ingriper i metabolismen och deras biokemiska struktur förändras inte av messenger RNA (mRNA) under läsningsprocessen. Det förklarar delvis DNA: s livslängd.
Halveringstiden för mitokondriellt DNA (mtDNA), under vilken hälften av de ursprungligen befintliga bindningarna mellan nukleobaserna sönderfaller, är starkt beroende av miljöförhållandena och varierar mellan cirka 520 år under genomsnittliga förhållanden med positiva temperaturer och upp till 150 000 år under permafrostförhållanden .
Som en del av RNA spelar nukleobaserna en något mer aktiv roll. I princip, när celler delar sig, bryts DNA-dubbelsträngarna upp och separeras från varandra för att kunna bilda en komplementär sträng, mRNA, som så att säga utgör arbetskopian av det genetiska materialet och fungerar som bas för valet och sekvensen av aminosyrorna från vilka avsedda proteiner monteras. En annan nukleisk bas, dihydrouracil, förekommer endast i det så kallade transport-RNA (tRNA), som används för att transportera aminosyror under proteinsyntes.
Vissa nukleobaser har en helt annan funktion som en del av enzymer som aktivt katalytiskt möjliggör och kontrollerar vissa biokemiska processer. Adenin uppfyller sin mest kända uppgift som en nukleotid i energibalansen i celler. Adenin spelar en viktig roll som elektrondonator som adenosindifosfat (ADP) och adenosintrifosfat (ATP) såväl som en komponent i nikotinamid-adenindinukleotiden (NAD).
Utbildning, förekomst, egenskaper och optimala värden
I den icke-fosforylerade formen består nukleobaser uteslutande av kol, väte och syre, ämnen som är allestädes närvarande och fritt tillgängliga. Kroppen kan därför syntetisera nukleobaser själv, men processen är komplex och energikrävande.
Därför föredras återvinning av nukleinsyror genom återvinning, t.ex. B. genom att bryta ner proteiner som innehåller vissa föreningar som kan isoleras och omvandlas till nukleinsyror med liten energiförbrukning eller till och med med energivinst. Nukleinsyror förekommer vanligtvis inte i ren form i kroppen, utan mestadels som nukleosider eller deoxynukleosider med en bifogad ribos- eller deoxiribosmolekyl. Som en komponent av DNA och RNA och som en komponent i vissa enzymer fosforyleras nukleinsyrorna eller deras nukleosider också reversibelt med en till tre fosfatgrupper (PO4-).
Det finns inget referensvärde för en optimal tillförsel av nukleobaser. En brist eller ett överskott av nukleobaser kan endast bestämmas indirekt via vissa störningar i ämnesomsättningen.
Sjukdomar och störningar
Den typ av faror, störningar och risker som bringas i samband med nukleobaserna är fel i antalet och sekvensen på DNA- eller RNA-strängarna, vilket leder till en förändring i kodningen för proteinsyntes.
Om kroppen inte kan avhjälpa felet genom sina reparationsmekanismer kommer det till syntesen av biologiskt inaktiva eller användbara proteiner, vilket i sin tur kan leda till milda till allvarliga metaboliska störningar. Det kan t.ex. B. genmutationer är närvarande som kan utlösa symptomatiska sjukdomar från början via metaboliska störningar, vilket kan vara obotligt. Men även i ett friskt genom kan kopieringsfel uppstå under replikering av DNA- och RNA-kedjorna, som påverkar metabolismen.
En känd metabolisk störning i purinbalansen är z. B. tillbaka till en genetisk defekt på x-kromosomen. På grund av den genetiska defekten kan inte purinbaserna hypoxantin och guanin återvinnas, vilket i slutändan främjar bildandet av urinstenar och gikt i lederna.


.jpg)