De komplexa cellulära och fysiologiska processerna i levande varelser kräver finjusterad reglering på molekylnivå för att säkerställa anpassningsförmågan hos ett djur eller en växt, till exempel till livsmiljön. För detta ändamål finns det många molekyler som ingriper i processer som cellkommunikation, metabolism eller celldelning. En av dessa molekyler är protein calmodulinsom med hjälp av kalcium påverkar funktionen hos många andra biologiskt aktiva proteiner.
Vad är calmodulin?
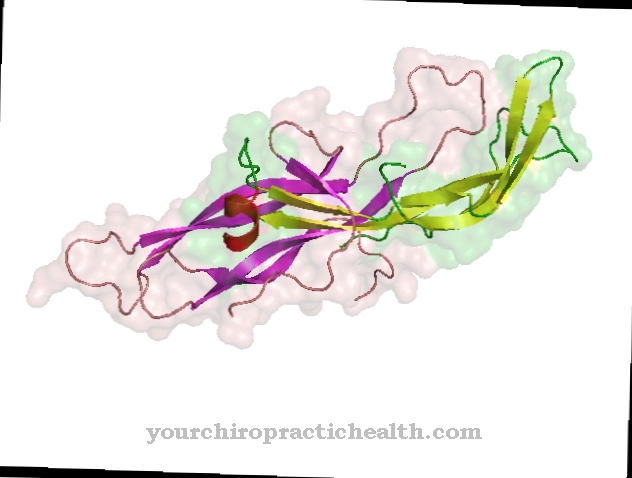
Calmodulin är ett intracellulärt reglerande protein som binder kalciumjoner. På grund av dess struktur tillhör den gruppen av EF-handproteiner. Formen på calmodulin, som består av 148 aminosyror och är 6,5 nm lång, liknar en hantel. Molekylmassan för denna proteinmolekyl är cirka 17 kDa.
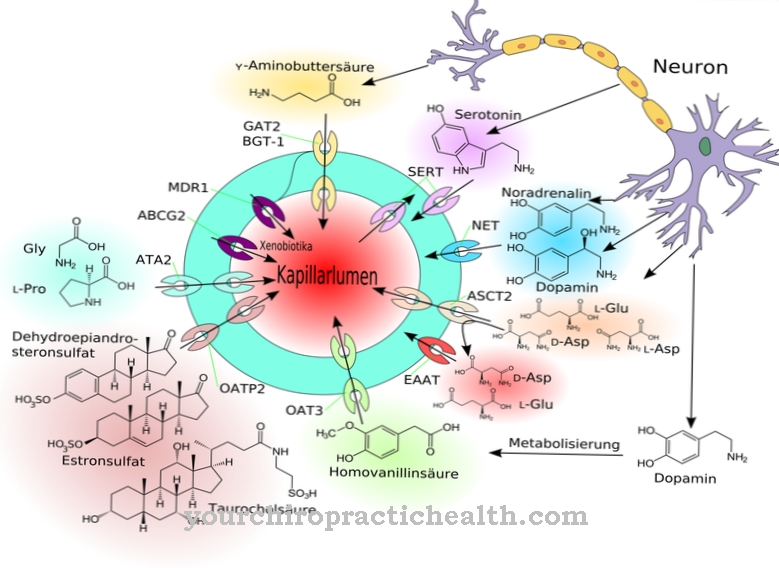
På grund av dess biologiska funktion i signalöverföring inom celler kan calmodulin också klassificeras som en andra messenger, dvs en sekundär messenger-substans som emellertid inte själv är enzymatisk aktiv. I de två sfäriska domänerna för proteinet finns det två spiral-loop-spiral-motiv på ett avstånd av 1,1 nm, till vilka totalt fyra kalciumjoner kan bindas. Denna struktur är känd som EF-handen. EF-handstrukturerna är förbundna med vätebindningar mellan de antiparallella beta-arken av calmodulin.
Funktion, effekt och uppgifter
Calmodulin behöver tre till fyra bundna kalciumjoner per molekyl för att vara aktiva. I det aktiverade tillståndet är det bildade kalcium-kalmodulinkomplexet involverat i regleringen av ett stort antal receptorer, enzymer och jonkanaler med en mängd olika funktioner. De reglerade enzymerna inkluderar fosfatas-calcineurin, som spelar en viktig roll för att reglera immunresponsen, och det endoteliala kväveoxidsyntaset (eNOS), som producerar NO, som bland annat används för avslappning av mjuka muskler och därmed för en utvidgning av Blodkärl.
Vid låga kalciumkoncentrationer aktiveras också adenylatcyklas (AC), vid höga kalciumkoncentrationer, å andra sidan, den enzymatiska motsvarigheten, fosfodiesteras (PDE). På detta sätt uppnås en kronologisk sekvens av regleringsmekanismerna: initialt sätter AC en signalväg i rörelse via produktionen av cyklisk AMP (cAMP), senare stängs detta igen av motståndaren PDE via cAMP-nedbrytning. Emellertid är den reglerande effekten av kalmodulin på proteinkinaser såsom CaM-kinas II eller myosin-lätta kedjekinas (MLCK), som kommer att förklaras mer detaljerat nedan, särskilt känd.
CAMKII kan binda en fosfatrest till olika proteiner och därmed påverka energimetabolismen, permeabiliteten för joner och frisättningen av neurotransmittorer från cellerna. CAMKII finns i särskilt höga koncentrationer i hjärnan, där den spelar en viktig roll i neuronal plasticitet, d.v.s. alla inlärningsprocesser. Men calmodulin är också nödvändigt för rörelseprocesser. I vila är koncentrationen av kalciumjoner i en muskelcell mycket låg och kalmodulin är därför inaktivt. Om muskelcellen är upphetsad, strömmar emellertid kalcium in i cellplasma och som en kofaktor upptar de fyra bindningsställena på kalmodulin.
Detta kan nu aktivera myosin-lätta kedjekinas, som förskjuter de kontraktila fibrerna i cellen och därmed möjliggör muskelkontraktion. Andra, mindre kända enzymer som påverkas av calmodulin är guanylatcyklas, Ca-Mg-ATPas och fosfolipas A2.
Utbildning, förekomst, egenskaper och optimala värden
Calmodulin förekommer i alla eukaryoter, som inkluderar alla växter, djur, svampar och gruppen av amoeboid varelser. Eftersom kalmodulinmolekylen i dessa organismer vanligtvis är strukturerad på ett relativt lika sätt kan det antas att det är ett evolutionärt mycket gammalt protein som växte fram i ett tidigt skede.
Som regel förekommer kalmodulin i relativt stora mängder i en cells plasma. I cytosolen i nervcellerna är till exempel den vanliga koncentrationen cirka 30-50 um, d.v.s. 0,03-0,05 mol / L Proteinet bildas under transkription och translation med användning av CALM-genen, av vilka det finns tre hittills kända alleler, som kallas CALM-1, CALM-2 och CALM-3.
Sjukdomar och störningar
Det finns vissa kemikalier som kan ha en hämmande effekt på calmodulin och är därför kända som calmodulin-hämmare. I de flesta fall är deras hämmande effekt baserad på det faktum att de transporterar kalcium ut ur cellen och på så sätt avlägsnar det från calmodulin, som då endast finns i det inaktiva tillståndet.
Dessa hämmande ämnen inkluderar till exempel W-7. Dessutom hämmar vissa psykotropa fenotiazinläkemedel calmodulin. Så breda som de reglerande funktionerna för calmodulin, så olika är de tänkbara defekterna och störningarna när proteinet inte längre kan aktiveras av kofaktorkalcium och de reglerade målenzymerna själva är mindre aktiva. Otillräcklig aktivering av CAMKII, till exempel, kan leda till en begränsning av den neurala plasticiteten, som utgör grunden för inlärningsprocesser.
Minskande aktivering av MLCK försvårar muskelsammandragningen, vilket kan leda till rörelsestörningar. Mindre aktivering av enzymet kalcineurin på grund av en kalmodulinbrist skulle påverka kroppens immunrespons och mindre aktivering av eNO: erna skulle leda till lägre NO-koncentrationer. Det senare orsakar huvudsakligen problem där kväveoxiden annars är tänkt att förhindra oönskad blodkoagulation och utvidga kärlen i syfte att förbättra blodcirkulationen. Det bör emellertid också nämnas vid denna punkt att kalciumsensorn Frequenin kan ta över de biologiska funktionerna för kalmodulin under vissa förhållanden och därmed ersätta molekylen.


.jpg)